
Fungi exhibit three major modes of
reproduction - vegetative, asexual and
sexual.Fungal

Fungi reproduce sexually and/or
asexually. Perfect fungi reproduce both sexually and asexually, while imperfect
fungi reproduce only asexually (by mitosis).
In
both sexual and asexual reproduction,
fungi produce spores that disperse from the parent organism by either floating
on the wind or hitching a ride on an animal. Fungal spores are smaller and
lighter than plant seeds. The giant puffball mushroom bursts open and releases
trillions of spores. The huge number of spores released increases the
likelihood of landing in an environment that will support growth.
Vegetative Reproduction
It is the type of reproduction which
involves the somatic portion of the fungal thallus. It occurs by the following
methods.
Fragmentation
In this process, the mycelium breaks
into two or more similar fragments either accidentally or due to some external
force. Each fragment grows into a new mycelium.
Budding
The parent cell produces one or more
projections called buds, which later develop necessary structures and
detach to grow into new individuals. Budding is common in unicellular forms
like yeast.
Fission
In this process, the parent cell
splits into two equal halves, each of which develop into a new individual.
Fission is also common in yeast.
Sclerotia
In some cases, as in Claviceps, the
hyphae become interwoven to form a compact mass and get surrounded by a hard
covering or rind. Such structures are called SCLEROTIA. They remain dormant
under unfavourable conditions and germinate into new mycelia on the return of
favourable conditions.
Rhizomorphs
In some higher fungi, several hyphae
may become interwoven to form rope-like structures called rhizomorphs. Under
favourable conditions, they resume growth to give rise to new mycelia.

Modes of Vegetative
Reproduction
Asexual Reproduction
Fungi reproduce asexually by
fragmentation, budding, or producing spores. Fragments of hyphae can grow new
colonies. Mycelial fragmentation occurs when a fungal mycelium separates
into pieces with each component growing into a separate mycelium. Somatic cells
in yeast form buds. During budding (a type of cytokinesis),
a bulge forms on the side of the cell, the nucleus divides mitotically, and the
bud ultimately detaches itself from the mother cell.
The most common mode of asexual
reproduction is through the formation of asexual spores, which are produced by
one parent only (through mitosis) and are genetically identical to that parent
(Figure 2). Spores allow fungi to expand their distribution and colonize new
environments. They may be released from the parent thallus, either
outside or within a special reproductive sac called a sporangium.
There are many types of asexual
spores. Conidiospores are unicellular or multicellular spores that are released
directly from the tip or side of the hypha. Other
asexual spores originate in the fragmentation of a hypha to form single cells
that are released as spores; some of these have a thick wall surrounding the
fragment. Yet others bud off the vegetative parent cell. Sporangiospores are
produced in a sporangium.
Zoospores
They are flagellated, motile spores
produced inside structures called zoosporangia. These spores do not have a cell
wall. Such spores are produced in lower fungi such as Achyla and Saprolegnia.
Sporagiospores
These are non-motile spores produced
inside structures called sporangia in fungi such as Rhizopus and Mucor. These
spores are dispersed by wind.



Modes of Asexual
Reproduction
Chlamydospores
These are thick walled resting
spores which arise directly from hyphal cells. They store reserve food.
Oidia
These are spore like structures
formed by the breaking up of hypha cells. They do not store reserve food and
hence cannot survive under unfavourable conditions. Such spores are produced in
Rhizopus.
Conidia
These are non-motile spores produced
singly or in chains at the tip of the hypha branches that are called
conidiophores. Such spores are produced in fungi like Aspergillus and
Penicillium.
|
Asexual reproduction
The ability to reproduce asexually
is common to almost all fungi.
1. Bud formation in
yeasts
In its simplest form asexual
reproduction is by budding or binary fission. The onset of the cellular
events is accompanied by the nuclear events of mitosis. If you have forgotten
these events please check in a basic text book.
The initial events of budding can
be seen as the development of a ring of chitin around the point where the bud
is about to appear. This reinforces and stabilizes the cell wall. Enzymatic
activity and turgor pressure the act to weaken and extrude the cell wall. New
cell wall material is incorporated during this phase. Cell contents are
forced into the progeny cell, and as the final phase of mitosis ends a cell
plate, the point at which a new cell wall will grow inwards from, forms.
|
Yeasts budding:

Separation of the bud from the
parent leaves a scar. When chains of yeast cells do not fully separated this
can create a pseudo-mycelium.
2. Fragmentation
Many fungi can reproduce by
fragmentation. Any mycelium that is fragmented or disrupted, provided that the
fragment contains the equivalent of the peripheral growth zone, can grow into a
new colony. Many fungi are sub-cultured using this hyphal fragment technique.
All of this weeks practical plates have been inoculated in this way with a cork
bore taken from a colonized donor plate. Cut mycelial tips do not regenerate,
but branches can form some distance from the damage point.
3. Sporulation
By far the most important type of
asexual reproduction is that of spore formation. Asexual reproduction is
extremely important to fungi. It is responsible for the production of large
numbers of spores throughout the year. These asexual spores are formed on a
phase of the fungal life cycle termed in some texts as the mitosporic, or
anamorphic phase. There can be more than one mitosporic state for each species
of fungus, and in some cases the mitosporic state of very different species can
look very similar. This has contributed to the problems of creating a taxonomy
for the fungi that only possess mitosporic states. The sexual stage of the
fungus can be termed the teleomorph, and the characteristics of this phase of
the life cycle are much more stable and reliable for taxonomic purposes.
The onset of asexual reproduction is
controlled by many different things. Some are environmental, like nutrient
levels, CO2 levels, light levels. Others can fungi have internal
time clocks and sporulate anyway in a preset part of the fungal life cycle
designed to spread and maximize colonization during one season.
Chytridiomycete asexual reproduction
Chytrids are quite distinct from
other fungi as they have extremely simple thalli and motile zoospores. Species
within this group are very simple in structure and may only consist of a single
cell, perhaps with rhizoids to anchor it on to a substrate.
Asexual reproduction in the chytrids
is by the production of motile zoospores, with a single, posterior flagellum,
in sporangia (Fig. 2). There will be a film of chytrid reproduction available
in the practical.

Figure
2. Holocarpic chytrid
This is a diagram of a holocarpic
chytrid, one where the entire thallus consists of only one cell with rhizoids.
These are usually parasitic on aquatic plants or fish. The fungus 'feeds' from
its substrate via its rhizoids. The entire cell contents will convert to motile
zoospores
Zygomycete Asexual Reproduction
Zygomycete fungal mycelium is coenocytic. At the onset of sporulation
large amounts of aerial hyphae are produced. The tips of these aerial hyphae
fill with cytoplasmic contents, and the nuclei undergo repeated mitosis. Around
each of the nuclei cytoplasm and organelles collect, and by the formation of
copious vesicles from the Golgi, each nucleus becomes isolated from the next by
a plasma membrane. Within the spaces created by this cytoplasmic cleavage,
spore walls begin to form, again by the fusion of Golgi vesicles containing
cell wall monomers and enzymes with the spore membrane. A sporangium forms. As
these events occur so there is considerable water uptake by the forming
sporangium, and as the columella forms the structure comes under considerable
turgor pressure. The large sporangia can contain up to 100,000 spores.
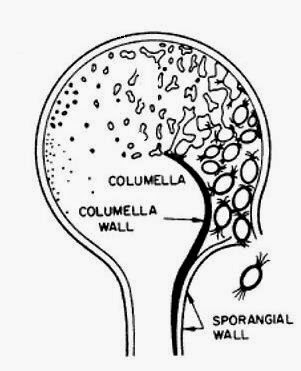
Fig.
3 Formation of a sporangium
This diagram is redrawn from
Brackers original and shows the development of a sporangium through time. As
nuclei undergo repeated mitosis so the Golgi produces membrane bound vesicles
filled with spore wall building materials. These coalesce around the nuclei to
form a spore. Eventually they are released.
Not all sporangia are as large as
this, there are many species with smaller, specialized sporangia, called
sporangiola, merosporangia and some are almost conidial, forming single spored
sporangia that are only distinguished from the conidium of the higher fungi by
the possession of a double spore wall. We will look at some of these structures
in the practical.
Asexual reproduction in the Ascomycetes (and deuteromycetes)
The process of spore formation in
most members of the higher fungal groups is again based largely on the
formation of aerial mycelium and the differentiation of the hyphal tip.
However, unlike the process seen in the Zygomycetes, the process here
involves something much more like the budding we see in the yeasts. This is
termed a blastic process, which involves the blowing out or blebbing of the
hyphal tip wall. The blastic process can involve all wall layers, or there can
be a new cell wall synthesized which is extruded from within the old wall. As
asexual reproduction is sometimes the only form of reproduction seen in some
fungi, we have in the past tried to construct elaborate taxonomic schemes based
on spore structure and production. However, as I said earlier, these features
are notoriously plastic and such schemes have largely been abandoned. The hypha
that creates the sporing (conidiating) tip can be very similar to the normal
hyphal tip, or it can be differentiated. The commonest differentiation is the
formation of a bottle shaped cell called a phialide, from which the spores are
produced. Phialide formation in the Ascomycete fungi:

Not all of these asexual structures
are single hyphae. In some groups the conidiophores (the structures that bear
the conidia) are aggregated. In the Moniliales all are single with the
exception of the aggregations termed coremia or synnema. These produce
structures rather like corn-stooks, with many conidia being produced in a mass
from the aggregated conidiophores.
Other species of Ascomycetes
and Deuteromycetes form their structures within plant tissue, either as
parasite and saprophytes. These fungi have evolved more complex asexual sporing
structures, probably influenced by the cultural conditions of plant tissue as a
substrate. These structures are called the sporodochium. This is a cushion of
conidiophores created from a psuedoparenchymatous stroma in plant tissue. The
pycnidium is a globose to flask-shaped parenchymatous structure, lined on its
inner wall with conidiophores. The acervulus is a flat saucer shaped bed of
conidiophores produced under a plant cuticle, which eventually erupt through
the cuticle for dispersal.

Sexual reproduction
Sexual reproduction with meiosis exists in
all fungal phyla (with the exception of the Glomeromycota).
It differs in many aspects from sexual reproduction in animals or plants.
Differences also exist between fungal groups and can be used to discriminate
species by morphological differences in sexual structures and reproductive
strategies.Mating experiments between fungal isolates may identify species on
the basis of biological species concepts. The major fungal groupings have
initially been delineated based on the morphology of their sexual structures
and spores; for example, the spore-containing structures, asci and basidia, can be used in the identification of ascomycetes and
basidiomycetes, respectively. Some species may allow mating only between
individuals of opposite mating type,
while others can mate and sexually reproduce with any other individual or
itself. Species of the former mating system
are called heterothallic, and of the latter homothallic.
Most fungi have both an haploid and diploid stage in
their life cycles. In sexually reproducing fungi, compatible individuals may
combine by fusing their hyphae together into an interconnected network; this
process, anastomosis, is required for the initiation of the sexual cycle.
Ascomycetes and basidiomycetes go through a dikaryotic
stage, in which the nuclei inherited from the two parents do not combine
immediately after cell fusion, but remain separate in the hyphal cells (see heterokaryosis).

In ascomycetes, dikaryotic hyphae of
the hymenium (the
spore-bearing tissue layer) form a characteristic hook at the hyphal
septum. During cell division, formation of the hook ensures proper distribution of the
newly divided nuclei into the apical and basal hyphal compartments. An ascus
(plural asci) is then formed, in which karyogamy
(nuclear fusion) occurs. Asci are embedded in an ascocarp, or
fruiting body. Karyogamy in the asci is followed immediately by meiosis and the
production of ascospores. After dispersal, the ascospores may germinate and form a
new haploid mycelium.Sexual reproduction in basidiomycetes is similar to that
of the ascomycetes. Compatible haploid hyphae fuse to produce a dikaryotic
mycelium. However, the dikaryotic phase is more extensive in the
basidiomycetes, often also present in the vegetatively growing mycelium. A
specialized anatomical structure, called a clamp connection,
is formed at each hyphal septum. As with the structurally similar hook in the
ascomycetes, the clamp connection in the basidiomycetes is required for
controlled transfer of nuclei during cell division, to maintain the dikaryotic
stage with two genetically different nuclei in each hyphal compartment. A basidiocarp
is formed in which club-like structures known as basidia generate
haploid basidiospores after karyogamy and meiosis. The most commonly known
basidiocarps are mushrooms, but they may also take other forms (see Morphology
section).
In glomeromycetes (formerly
zygomycetes), haploid hyphae of two individuals fuse, forming a gametangium,
a specialized cell structure that becomes a fertile gamete-producing cell. The gametangium develops into a zygospore,
a thick-walled spore formed by the union of gametes. When the zygospore
germinates, it undergoes meiosis, generating new haploid hyphae, which may then form asexual
sporangiospores. These sporangiospores allow the fungus to rapidly disperse
and germinate into new genetically identical haploid fungal mycelia.Sexual
fusion in fungi is of different types, as follows :
Planogametic Copulation
Here motile gametes called planogametes
undergo fusion. When both the gametes are motile and morphologically similar,
the fusion process is called isogamy.
Eg.: Synchytrium When both the
gametes are motile but differ in their size, the fusion process is called anisogamy.
Eg.: Allomyces. When one gamete
(male) is smaller and motile and the other (female) gamete is larger and non
motile, the fusion process is called heterogamy.
Gametangial Contact
Here, gamete bearing structures
called gametangia come closer to each other and develop a fertilisation tube
through which the male gamete migrates into the female gametangium.
Eg. : Phytophthora, Albugo.
Gametangial Copulation
Here,
the gametangia fuse with each other, lose their identity and develop into a
zygospore
Eg.:
Mucor, Rhizopus
Spermatisation
In some fungi like Puccinia, tiny
unicellular spore like structures called spermatia are formed. They get
transferred to female gametangia through various agencies.



Types of Sexual
Reproduction in Fungi
Somatogamy
In examples like Agaricus, fusion
occurs between two somatic cells and involves only plasmogamy. This results in
the formation of dikaryotic hyphae. Hence, the process is called
dikaryotization.
Homothallism And Heterothallism
Based on the compatibility in sexual
reproduction the fungal hyphae can be distinguished into two types homothallic
and heterothallic. In homothallic forms, fusion occurs between the
genetically similar strains or mating types. In such forms,
meiosis results in the formation of genetically identical spores. In the
heterothallic forms, fusion occurs between the genetically different mating
types or strains. The strains are genetically compatible and are designated
as + strain and strain. In such forms meiosis results in the
formation of both the strains, in equal numbers. Heterothallism is a device to
prevent inbreeding and promote out breeding.
Sexual reproduction
Sexual reproduction introduces the
possibility of variation into a population, and this is why most fungi have a
sexual phase. To achieve sexual reproduction it is necessary to have two mating
type haploid nuclei (n + n), or a diploid (2n) nucleus. In the case of the two
haploid nuclei they must fuse to form a diploid first, but once fused the
nuclei undergo meiosis, which is the reduction division that potentially brings
about variation in the progeny. These event are followed by the formation of
spores, which in most cases are resting spores that can withstand adverse
conditions.
Sexual reproduction in
the chytridiomycetes:
Sexual reproduction occurs in some
members of the chytrids by the production of diploid spores after gametic or
somatic fusion of two different mating types. The resulting spore may germinate
to produce a diploid vegetative mycelium or it may undergo meiosis to produce a
haploid mycelium. The diploid mycelium can also produces resting sporangia in
which meiosis occurs, generating haploid zoospores that germinate to produce
haploid vegetative mycelium:

Sexual reproduction in
the Zygomycetes:
There are two possible nuclear
states in the mycelia of this group of fungi. They can have a single type of
nucleus in their mycelium, a condition termed termed homothallism, or they can
contain the two mating type nuclei within their mycelium, termed
heterothallism. If the fungus is homothallic the first event in the onset of
sexual reproduction has to be somatic fusion. This is termed conjugation. To
achieve such a mating it is necessary to attract each other and an elaborate
sequence of cellular and biochemical events have been established for some of
these fungi. This signalling involves the secretion of inducer molecules that
are responsible for causing the formation of zygophores, modified hyphal tips,
and these then grow towards each other long a gradient of hormone. The exact
sequence is shown below. Sexual reproduction in the Zygomycetes:

Once in contact the two zygophores
fuse, and then the nuclei fuse to form the diploid. Meiosis occurs, producing
four haploid nuclei, but three may degenerate. The timing of fusion varies from
species to species.
Sexual reproduction in
the Ascomycetes:
In this group of fungi there are no specialized
organs of hyphal fusion, different mating type mycelia merely fuse with each
other to form transient dikaryons, mycelia with two mating type nuclei within
it. The dikaryotic mycelium can differentiate to from varying amounts of
sterile mycelium around what is to become the fertile tissue of the fruit body.
In yeasts, a single, diploid yeast will undergo meiosis, producing four haploid
progeny cells, but in more complex fungi there are a sequence of cellular and
nucleic events that ensure an organized fertile layer.

Spores are delineated around these
nuclei in a process called free cell formation, and as most of the cytoplasm is
contained around the nucleus and within the spore wall, all that is left
outside is cell sap. These modified hyphae are termed Asci, and the spores that
are held within them are termed ascospores. The asci are often found packed
tightly with other asci, and between a dense layer of supporting sterile
tissue. Often the structure is large enough to be seen with the naked eye.

The asci can be aggregated together
in various sorts of fruit body which we will see in the practical, including
the, cup fungi (Discomycetes, apothecial), the flask fungi,
(Pyrenomycetes, perithecial), the mildews (Plectomycetes cleistothecial)
and the fungi with black, crusty stromata (Loculoascomycetes,
pseudothecial fungi). There are also the yeasts, Hemiascomycetes,. Their
ascospores are normally formed in loose asci and are not actively discharged.
We have not looked at these. When they form ascospores in fruit pulps or
liquids they are usually liberated by the disintegration of the ascus wall.
Sexual reproduction in
the Basidiomycetes:
Basidiomycetes are characterised by the most complex and large structures
found in the fungi. They are very rarely produce asexual spores. Much of their
life cycle is spent as vegetative mycelium, exploiting complex substrates.
A preliminary requisite for the
onset of sexual reproduction is the acquisition of two mating types of nuclei
by the fusion of compatible mycelium. This creates a dikaryon where single
copies of the two mating type nuclei are held within every hyphal compartment
for extended periods of time. Maintenance of the dikaryon requires elaborate
septum formation (clamp connections) during growth and nuclear
division.

Onset of sexual spore formation is
triggered by environmental conditions and in the larger Basidiomycetes begins
with the formation of a fruit body primordium. The primordium expands and
differentiates to form the large fruit bodies of mushrooms and toadstools. The
mycelium within this structure remains as a dikaryon, diploid formation only
occurring within the modified hyphal tip called the basidium. Meiosis occurs within
the basidium, and the four products are extruded from the tip of the basidium
on sterigma (below). Usually this event occurs across a large area of basidia
called a hymenium, or fertile layer. It is usually formed over an extensive
sterile layer of tissue like a mushroom gill.

There are three major divisions in
the basidiomycetes.
- Hymenomycetes. Basidia are in extensive fertile layer which are susceptible to rain when exposed. Spores are actively discharged from a protected hymenium when ripe. This group includes mushrooms and toadstools, boletes, brackets and coral fungi.
- Gasteromycetes. Hymenia line closed cavities in an initially closed fruit body. Basidiospores are released passively by autolysis of the hymenium, and basidiocarps disintegrate at maturity. This group includes earth balls, puff-balls, stinkhorns and birds nest fungi.
- Teliomycetes. These are the rusts and smuts, neither of which form large, conspicuous, fruit bodies but invade plants and produce characteristic sporulating lesions in plant tissue.
Spore dispersal and the air spora
There are two major functions of
fungal spores, dispersal and survival. Often these two requirements are met by
two different spores formed at different points in a fungus’ life cycle. Some
are survival spores formed in response to adverse abiotic conditions that can
include desiccation, high UV, high/low temperatures or starvation. Biotic
factors can also induce sporulation including competition, antagonism, and
pathogens presence. These spores have thick cell walls, and lots of reserves.
Dissemination spores are spores that
are smaller, with thin cell walls, and limited reserves, and will germinate
readily when on a suitable substrate. They are formed as part of the active
life cycle of the fungus and are often concerned with epidemic spread of a
pathogenic species from plant to plant, or with rapid colonization of a
substrate.
Spores in general:
By wet weight spores generally
contain 25% protein and 20% fat, and they have a low water content relative to
vegetative mycelium. Cell walls of spores are generally not fibrillar, but they
are multi-layered and often contain melanin and have ornamentations.
Spores contain all normal mycelial
organelles. Respiratory reserves include lipids, glycogen, phospholipids and
polysaccharides that can include sugar alcohols like Trehalose). Respiration
rates in spores are only 1-4% those of vegetative mycelium, but obviously the
more reserves a spore has, the longer it will survive.
Dormancy:
Dormancy occurs when spores do not
immediately germinate after formation. Dormancy is a break in the life cycle.
There are two types, endogenous (constitutive) and exogenous (induced).
Endogenous dormancy is due to some internal quality of the spore, a barrier to
water or nutrient entry, a metabolic block, or an inhibitor. Self inhibition
prevents spores from germinating in dense suspensions. It can be by excessive
sensitivity to oxygen or carbon dioxide levels, nutrient competition, or most
usually due to the presence of inhibitors. These molecules are often active in
the 1-10 nanomolar range. These inhibitors have to be leached away before
germination takes place.
There can also be physical barriers
to germination. In one of the athlete foot fungi, Microsporium gypseum,
there is a protein layer around the spore which prevents the uptake of water.
This layer is removed by the action of a fungal acid phosphatase enzyme. This
enzyme is inhibited by high levels of phosphate, and until phosphate levels in
the environment drop the fungus spore does not germinate.
Endogenous (induced) dormancy occurs
because of some external condition, and whilst these conditions prevail the
spore will not germinate. As soon as the limiting factor is removed the spore
germinates.
Table
1. A summary of the characteristics of fungal spores with endogenous and
exogenous dormancy:
|
Endogenous
dormancy:
|
Exogenous
dormancy:
|
|
Displaced from point of origin
|
Remain at point of origin
|
|
Definite launch mechanism
|
Released by autolysis
|
|
Small and thin walled spores
|
Large and thick walled
|
|
Short survival time
|
Survive for a long time
|
|
Germinate readily under suitable
conditions
|
Germinate after a specific
stimulus or removal of an inhibitor
|
Optimal environmental signals
trigger the end of dormancy and the onset of germination. Chemical stimuli can
trigger germination. This is frequently seen in pathogens where host compounds
can act as germination stimulants.
Germination begins with imbibition,
the uptake of water, which can cause a 3 to 20 fold increase in size. Spherical
growth also accounts for some of the swelling. Eventually polarized growth
starts, with the emergence of a germ tube from the spore. The spore wall may be
ruptured and a new cell wall covered germ tube emerges, or the spore wall may
be softened and the germ tube then emerges.
Air spora:
In between formation of spores and
their eventual germination is a phase where spores are disseminated from their
point of origin. Many fungi have elaborate mechanisms for getting their spores
into the atmosphere which the best medium through which to spread spores. Many
spores are very dry and friable, which means they are light enough to be lifted
by air currents into the turbulent air above the boundary layer. Others have
active spore guns that fire spores up into the atmosphere.
The end result is that over both
countryside and towns there is a characteristic air spora. In the countryside
this is very typically full of spores from pathogens of agricultural crops,
from saprophytes of plant structures and from decaying matter. Species like Penicillium
and Cladosporium tend to predominate. In the towns there are fewer
agricultural pathogens but there are still hundreds of spores per cubic litre
of air. Within homes and workplaces spore numbers can be even higher, and the
species distribution tends to differ. In warm, dry areas Aspergillus
spp. can become significant members of the air spora.
In normal circumstances these fungal
spores pose little or no hazard, our immune systems have evolved with these
spores and we are at no risk from them. However, in a population that contains
a significant number of individuals that are for whatever reason
immuno-compromised, these spores can represent a hazard that as yet has not
been quantified. Furthermore, there is increasing evidence to show that
inhalation of fungal spores can cause significant allergic responses in atopic
individuals. This is an area of very active research.
0 comments:
Post a Comment